Eliökunnan luokittelu on aina ottanut uusia harppauksia tutkimusmenetelmien kehittymisen myötä. Ensimmäisiin tieteellisiin luokitteluihin päädyttiin rakenteellisten tuntomerkkien avulla.
Eliöiden rakenteita verrattiin ja samankaltaisimpien eliöiden oletettiin olevan läheisintä sukua toisilleen. Aluksi tutkimusvälineinä oli suurennuslaseja ja valomikroskooppeja, myöhemmin sukulaisuussuhteita pystyttiin tarkentamaan elektronimikroskopian avulla. Tumattomia eliöitä on luokiteltu ulkonäön lisäksi biokemiallisten ominaisuuksien, kasvuolosuhteiden ja taudinaiheutuskyvyn perusteella, sillä niissä on hyvin vähän mikroskoopillakaan nähtäviä ulkonäkötuntomerkkejä. Keskeinen edistysaskel eliökunnan rakenteen ymmärtämisessä saatiin, kun luokittelussa otettiin käyttöön DNA-tuntomerkit. Ne ovat tarjonneet valtavan aineiston evoluutiohistorian ja systematiikan tutkimukseen.
Parhaillaan eliökunnan luokittelu on muutoksen alla, sillä uusia eliöitä ja kokonaisia eliöryhmiä löydetään ja voidaan tutkia DNA-aineiston pohjalta. Yksittäisten geenialueiden vertailun lisäksi nykyään pystytään analysoimaan eliöiden sukulaisuussuhteita kokonaisten genomien aineistoilla, mikä muuttaa puolestaan ymmärrystä jo aiemmin tunnettujen eliöiden suhteista toisiinsa. DNA-tuntomerkkien avulla pystytään myös selvittämään rakenteiltaan ja toiminnoiltaan hyvin erilaisten eliöryhmien suhteita toisiinsa.
Erillistä tumattomien ryhmää ei ole
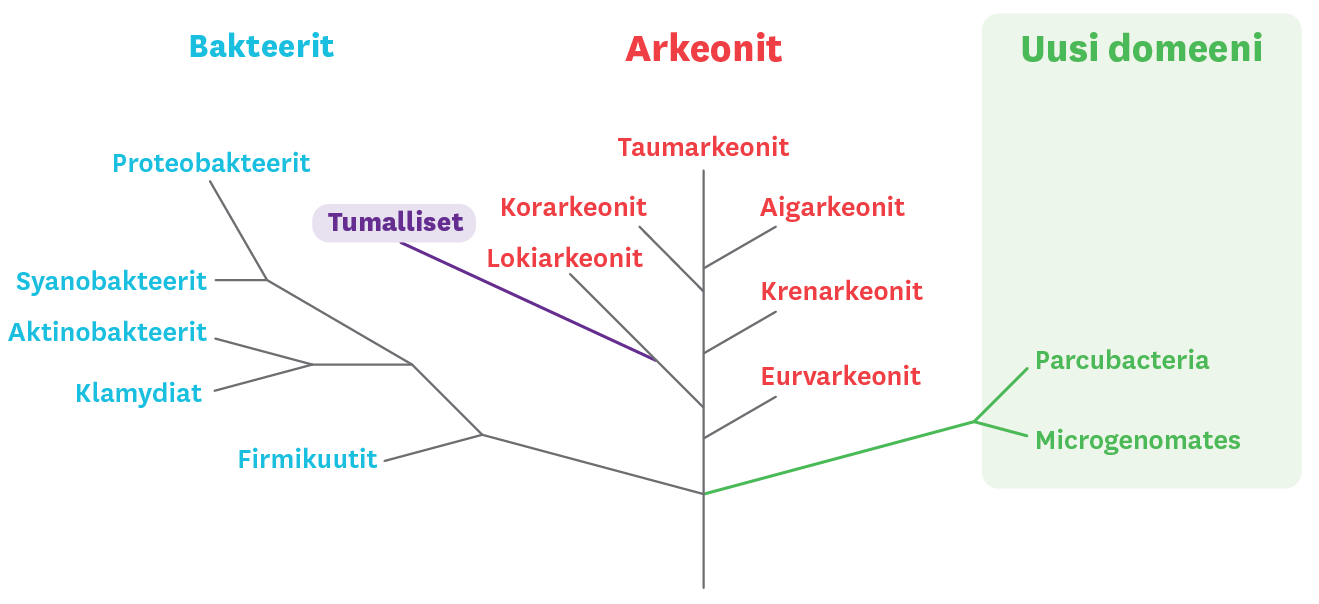
1990-luvulta on hyväksytty laajasti eliökunnan jako kolmeen domeeniin: bakteereihin, arkeoneihin ja tumallisiin. Jo parikymmentä vuotta on geneettisen aineiston perusteella ollut selvää, että tumalliset ja arkeonit ovat läheisempää sukua toisilleen kuin kumpikaan ryhmä on bakteereille. Arkeoneja ja tumallisia pidettiin kuitenkin erillisinä ryhminä, joilla on ollut yhteinen esi-isä. Tämä käsitys muuttui, kun vuonna 2015 löydettiin Lokiarkeonien ryhmä ja voitiin osoittaa, että tumalliset ovat kehittyneet suoraan Lokiarkeoneista. Näin tumallisista on tullut osa arkeonien sukuhaaraa. Myös biokemialliset havainnot tukevat tätä luokittelua. Naturan numerossa 3/2017 Timonen ja Rinta-Kanto kertovat lisää arkeoneista ja tumallisten evoluutiohistoriasta.
Tumallisten ja arkeonien sukuhaarojen yhdistyessä eliökunta jakautuisi siis karkeasti kahteen domeeniin, bakteereihin ja arkeoneihin. Kolmijaosta ei näillä näkymin kuitenkaan tarvitse luopua. DNA-menetelmien avulla on muodostumassa uusi sekä bakteereista että arkeoneista poikkeava eliöryhmä, johon kuuluu aiemmin bakteereihin luetut ryhmät Parcubacteria ja Microgenomates. Tämän ryhmän vakiintuminen osaksi eliökunnan hyväksyttyä rakennetta saattaa tosin viedä aikaa, sillä eliöt ovat äärimmäisen pieniä eikä niistä toistaiseksi ole puhdasviljelmiä.
Virusten määrittely vaikeaa
Myös virukset saattavat muuttaa eliökunnan perusjakoa, jos ne hyväksytään osaksi eliökuntaa. Viruksia ei ole perinteisesti luettu osaksi eliökuntaa, koska ne eivät tee solun ulkopuolella mitään tai lisäänny itsenäisesti. 2000-luvulla on löytynyt uusia varsin suurikokoisia viruksia, joilla on myös poikkeuksellisen suuri genomi ja sellaisia geenejä, joita virusten ei pitäisi tarvita toimintoihinsa. Jättiläisvirukset ovat saaneet nimekseen muun muuassa mimivirus ja megavirus. Osa tutkijoista tulkitseekin nykyään jättiläisvirukset omaksi domeenikseen. Yhtenä keskeisenä perusteena virusten lukemiseksi eliökuntaan on pidetty sitä, että virukset sisältävät nukleiinihappoja. Äärimmillään eliökuntaan on ehdotettu viidettäkin domeenia, prioneja, jotka ovat proteiinin kaltaisia hiukkasia. Prionit eivät sisällä nukleiinihappoja eli DNA:ta tai RNA:ta.
Eliökunnan monimuotoisuus metagenomiikan perusteella
Viime aikoina käsitystä eliöiden monimuotoisuudesta on ravistellut tunnistettujen tumattomien eliöryhmien määrä. Erilaisia bakteereita ja arkeoneja on löydetty huikeita määriä kun erilaisten ympäristöjen DNA:ta on monistettu ja tutkittu eliöiden esiintymistä suoraan niiden sisältämän eri eliöistä peräisin olevan DNA:n perusteella. Aiemmin eliöistä tarvittiin yksinomaan tätä eliötä sisältävä näyte, jotta DNA-koodi voitiin selvittää. Koska bakteerit ja arkeonit useimmiten elävät yhteisöinä, eikä niitä kokonsa vuoksi voi poimia yksittäiskappaleina, on niiden tutkiminen ollut aiemmin vaikeaa tai mahdotonta. Viime vuonna julkaistussa suoraan DNA-analyysiin eli metagenomiikkaan perustuvassa eliökunnan kuvauksessa näkyi 92 nimettyä bakteerien pääjaksoa, 26 arkeonipääjaksoa ja viisi tumallisten ylätason ryhmää. Ainakin osa tumattomien pääjaksoista saanee sukulaisuussuhteisiin perustuvan luokittelun vakiintuessa kuntatason statuksen. Valtaosa bakteerien ja arkeonien ryhmistä on toistaiseksi vailla puhdasviljelmäksi eristettyä ryhmän edustajaa ja lajitason taksonomian kuvaaminen on kesken.
Tumallisten tapaus
Käsitys tumallisten luokittelusta on ottanut aimo harppauksia genomiikan aikakaudella. Aiemmin käytetyissä yksittäisissä geenialueissa, (lähinnä 16S ribosomaalinen RNA) ei ole ollut riittävästi erotuskykyä tumallisten ryhmien sukulaisuussuhteiden selvittämiseksi luotettavasti. Useisiin geenialueisiin perustuvat tutkimukset osoittavat tällä hetkellä, että tumalliset jakautuvat viiteen ylätason ryhmään, joista tutkijakunta on melko yksimielinen: peräsiimaiset (Opisthokonta), amebat (Amoebozoa), kasvit (Archaeplastida), kromalveolaatit (Chromalveolata) ja ekskavaatit (Excavata). Peräsiimaisista eliöistä meille ovat tutuimpia eläimet ja sienet. Ameboihin puolestaan kuuluvat bakteereilla herkuttelevat kirkasväriset limakot ja erilaiset kuorelliset ja kuorettomat amebat. Kasveja ovat hyvin tuntemiemme vihreiden kasvien lisäksi myös punalevät ja pikkuiset glaukofyytit. Kromalveolaattien monimuotoisen ryhmän eliöt elävät usein erilaisissa yhteisöissä ja ne suosivat monenlaista yhteistyötä toistensa ja muiden eliöiden kanssa. Ekskavaateista löytyy enimmäkseen erilaisia yksisoluisia, joista monet ovat loisia. Ihmisille tuttuja ovat esimerkiksi unitautiloisio ja Giardia.
Oppimateriaaleissa käytetään vielä yleisesti luokittelua kasveihin, sieniin, eläimiin ja alkueliöihin, mutta tämä luokittelu ei vastaa eliöiden sukulaisuussuhteita ja siitä tulisi luopua. Aika näyttää tuleeko näistä viidestä tumallisten ylätason ryhmästä uudet kunnat vai nimetäänkö näitä varten jokin uusi taksonominen taso kuten yläkunta. Tällöin esimerkiksi eläinkunta saisi pitää kuntatason nimensä, mutta tumallisten kuntia olisi yhteensä vähintään kolmekymmentä. Lisäksi lokiarkeonit, joihin peräsiimaiset sisältyvät, tarvitsisivat vielä yläkuntaakin ylemmän statuksen.
Käytäntöjä tarvitaan
Bakteerien ja arkeonien valtavan monimuotoisuuden kuvaaminen asettaa haasteita luokittelun ja nimeämisen periaatteille. Bakteerien ja arkeonien uusien suurryhmien kuvaamiselle ei ole vakiintuneita käytäntöjä. Uusia pääjaksoja on karkeasti ottaen kuvattu, kun ryhmä ei ole yleisimmin käytetyn geenialueen 16S rRNA:n perusteella ollut selkeä osa aiemmin kuvattua pääjaksoa. Eläinten ja kasvien luokittelussa yleisessä käytössä oleva periaate on, että kuvausten tulisi aina perustua useamman geenialueen vertailuun. Lisäksi pitäisi nimetä ainoastaan yhden kantamuodon jälkeläisistä koostuvia ryhmiä. On myös ehdotettu, että pääjaksosta tulisi pystyä tunnistamaan ryhmälle yhteisiä piirteitä ja että ainakin yksi pääjakson eliö tulisi olla eristetty puhdasviljelmäksi. Kaiken kaikkiaan on toivottu malttia ja riittävän, kokonaisiin genomeihin perustuvaa aineiston, kertymistä. Eliölle sopivan nimen valinta ei sekään ole yksinkertaista. Historiallisista syistä eläinten, kasvien (myös jäkälien ja sienten), virusten ja bakteerien (myös arkeonien) nimeämissäännöstöt ovat erilaiset ja mikään niistä ei määrittele luokkaa korkeamman ylätason ryhmän oikean nimen valintaa.
Lajikäsitys uusiutuu
Myös lajikäsitys asettaa haasteita, kun kohteena on koko eliökunta. Kun selkärankaisten lajirajaukseen on perinteisesti liitetty lisääntymiskykyisten jälkeläisten tuottaminen ja yleensä myös rakenteellisia lajituntomerkkejä, perustuvat bakteerien lajit pääosin geenialueiden samankaltaisuuteen. Tällä hetkellä rajanveto eri lajien välillä tehdään hyvin eri perustein eri eliöryhmissä. Esimerkiksi bakteerien DNA-erojen täytyy olla huomattavasti suuremmat kuin vaikkapa eläinten, jotta voidaan perustella uuden lajin erottaminen muista. Lajien välillä tapahtuu myös paljon geenienvaihtoa. Useimpien ellei jopa kaikkien lajien perimät ovat eri alkuperistä tulleen DNA:n mosaiikkeja. Esimerkiksi tumallisten DNA:n monistamiseen tarvittavat geenit ovat peräisin arkeoneilta mutta energia-aineenvaihduntaan tarvittavista geeneistä suuri osa bakteereilta. Eri geenilinjoja seuraamalla sukupuu siis rakentuu erilaiseksi.
Aineiston käsittely on haastavaa
Genomien rakennetutkimus ja evolutiivinen kehitysbiologia tuottavat valtavasti lisäaineistoa eliökunnan sukulaisuussuhteiden tutkimukselle. Sen suurin haaste ei monesti olekaan aineiston puute. Merkittävä käytännön ongelma on miten käsitellä aineistoa. Eliöiden sukulaisuussuhteiden ratkaiseminen kuuluu laskennallisesti äärimmäisen vaikeiden ongelmien piiriin. Tähän mennessä ei ole pystytty kehittämään menetelmää, joka ratkaisee tällaisen ongelman varmasti oikein ja sellaisetkin haut, jotka pyrkivät hakemaan todennäköisimpiä vaihtoehtoja, vaativat tavattoman suurta laskentakapasiteettia.
Toinen keskeinen ongelma on vertailukohdan puuttuminen. Kaikki evolutiivisia uutuuksia etsivät analyysit hakevat ratkaisun niin kutsutun ulkoryhmän avulla. Menetelmät perustuvat evoluutioteoriaan ja tulos lasketaan suhteessa tiettyyn aiemmin kehittyneeseen eliöön. Tutkimuksen kohteena olevien eliöiden ominaisuuksien muutokset verrattuna ulkoryhmän ominaisuuksiin katsotaan analyysissä evolutiivisiksi uutuuksiksi, joiden avulla tutkittavia eliöitä sitten luokitellaan. Ulkoryhmää ei maapallon koko nykyistä eliöstöä tutkittaessa ole olemassa. Ainoaksi mahdollisuudeksi jää päätellä oletettavan ulkoryhmän tuntomerkit mahdollisimman hyvin perustein.
Digitaaliset oppimisympäristöt avuksi
Eliökunnan monimuotoisuuden kuvittaminen on vaativaa, erityisesti, jos niissä pyritään kuvaamaan kehityslinjojen suhteita täsmällisesti, mutta silti luomaan selkeitä kuvia. Luonnollisesti sukulaisuussuhteissa on vielä paljon tarkennettavaakin. Digitaaliset oppimisympäristöt tarjoavat monipuolisen mahdollisuuden monimuotoisuuden kuvaamiseen. Yksi esimerkki löytyy osoitteesta: http://www.onezoom.org/life/. Hanketta on kuvattu esimerkiksi tässä videossa https://www.youtube.com/watch?v=LZ3n3mV4uVc (englanninkielinen).
Teksti:
Aino Juslén, eläintieteen yksikön johtaja, Luonnontieteellinen keskusmuseo Luomus
Jyrki Muona, professori, Luonnontieteellinen keskusmuseo Luomus
Sari Timonen, yliopiston lehtori, Mikrobiologian osasto, Helsingin yliopisto
